 |
| |||||
|
| ||||||||
|
| ||||||||
Home  Publications Publications |
|
|
Print Friendly Nature 367:637-639 Link to companion
News and Views article by R.W. Guillery Other than the elimination of the optic chiasm, no other midline
abnormalities are evident in mutant dogs. In seven of eight cses, the
crossed component of the optic chiasm is absent (Fig 1B). The optic
nerves fail to approach midline, and maintain a separation of 5 to 8 mm
as they enter the thalamus and midbrain. In one animal (M2), a small
contralateral retinal projection was apparent (Hogan and Williams,
1995). The shape and lamination pattern of the LGN in the normal dog
resembles that of other carnivores. In comparison to the cat, the
central field representatin in the dog is positioned more rostrally, and
the caudal, vertically oriented limb of the nucleus is more prominent (Figu
2a) (1,6). The LGN of the mutant dogs closely resembles that of normal
dogs (Fig 2b). A prominent interlaminar zone separates layers A and A1
through the central part of the nucleus. In contrast to normal dogs, the
interlaminar zone does not extend to the rostromedial border of the
nucleus. In this area, which represents central visual fields, layers A
and A1 are fused (Fig. 2b, c). Upper visual fields are located
caudomedially, and lower visual fields are located rostrolaterally in
the LGN of both normal and mutant dogs. As in other carnivores (7),
receptive fields recorded from neighbouring segments of layers A and A1
in normal dogs view the same area of contralateral visual space. In mutants, as in normal dogs, visual stimuli in the contralateral
visual field evoked vigorous responses in layer A1 (Fig. 3). This
relatively small, normal ipsilateral temporal projection is unperturbed
by the aberrant ipsilateral input of nasal retina to neighbouring layer
A (Figs 3, 4). In each mutant, the entire aberrant nasal projection
establishes functional connections throughout ipsilateral layer A. These
aberrant ipsilateral fields extended to 90-100 degrees of azimuth and
included the region occupied by the contralateral monocular segment of
layer A in normal dogs. The misconnection of the nasal retina leads to a
severe misalignment of visual maps in layers A and A1. Within a large
central segment of the mutant LGN, we repeatedly recorded abrupt
reversals in receptive field azimuth (Fig. 3, curved arrows). The size
of these reversals varied systematically with the mediolateral position
of the penetration. In the rostral and medial part of the LGN, small
jumps in azimuth were observed. In the caudal and lateral LGN, jumps in
azimuth of as much as 35 degrees were encountered (Fig. 3). These
reversals across the vertical meridian representations were later
correlated with the border between A and A1. The achiasmatic Belgian sheepdog is the first mutant vertebrate in
which the size of the ipsilateral retinal projection is increased.
Interestingly, achiasmatic humans have also been described recently (8).
The achiasmatic mutation has an effect opposite to that produced by
albinism, which increases the size of the contralateral projection.
However, the achiasmatic mutation is not a simple anti-albino. In
albinism, the line of decussation is shifted toward the temporal edge of
the retina and there is almost invariably a residual uncrossed
projection from the far temporal retina (9,13). In contrast, in
achiasmatic dogs, the phenotype is more extreme and there is usually no
residual crossed projection. Although the misrouting of axons in mutant
dogs has an opposite patterns to that seen in albinos, the fundamental
mapping errors are reminiscent of those observd in the LGN of Siamese
cats and albino ferrets (14-17). The novel pattern of misrouting of retinal fibers in achiasmatic
mutants provides a challenging natural experiment that can be used to
study processes responsible for generating visual maps and LGN
lamination (Fig. 4). In the achiasmatic dog, a single retina innervates
a structure that normally receives input from two retinae. A single
input that projects to a single nucleus might be expected to produce a
single map in an unlaminated nucleus. This is a pattern found in the LGN
after unilateral enucleation early in development (18-20) (Fig. 4d) and
also found normally in another retinal target—the superior colliculus
(21). In fact, the LGN of mutants is laminated and nasal reintal fibres
terminate in ipsilateral layer A with the same anatomical polarity
normally seen in the contralateral LGN—the central nasalpart of retina
projects into the rostral and medial part of the LGN and the peripheral
nasal part of the retina projects inot the caudal and lateral part of
the nucleus. As these fibres have not corssed midline, the physiological
poloarity of the projection is reversed, both with respect to that in
normal dogs and with respect to that in the relatively normal layer A1
of mutant dogs. This results in mirror-image maps in layers A and A1
that cover the entire visual space and which are congruent only at the
representation of the vertical meridian (Fig. 4b). It is possible that
the vertical meridian is duplicated in A and A1 and that thesee layers
contain independent, but fused map. It is also possible that there is a
single map wrapped around on itself at the vertical meridian
representation. The well-ordered mirror-image maps in the mutant LGN
indicate that genes that control the crossing of retinal axons at the
chiasm differ from those that regulate the formation of maps in the LGN.
Despite its monocular innervation, three relatively normal
characteristics of the mutant LGN are surprising. It is not obvious why
the so-called binocular segment of layers A and A1 would be preserved in
mutants (Fig. 2), why nearly symmetrical receptive field positions would
be recorded across the A/A1 border (Fig. 3), or why the point at which
the layers are fused would correspond to the vertical meridian. All
three features can be explained if positional specificity between
retinal axons and LGN cells is rigidly preserved, despite the misrouting
of nasal axons at the chiasm. The preservation of target recognition in
mutants most probably reflects a fixed chemoaffinity between subsets of
retinal axons and LGN neurons (22, 24). This mutation shows that the
selection of a target site in the LGN is not controlled by the eye of
origin (namely, ipsilateral or contralateral), but rather that the
criticial factors are the retinal position from which axons originate
and positional specifity within the LGN. ACKNOWLEDGEMENTS. This work was supported by NIH and the National Eye
Institute. Our thanks to A. de LaHunta and J. Cummings and M. Young for
their help in starting this project. We thank B. Belton and T. Mandrell
for animal care; T. Kimble for technical help; J. Kaas for sharing his
facilities at Vanderbilt University with us, and N. Berman, E. Geisert,
L. Dell'Osso, and D. Goldowitz for discussion. References Kaas, J. H., Guillery, R. W. & Allman, J. M. Brain Behav. Evol 6,
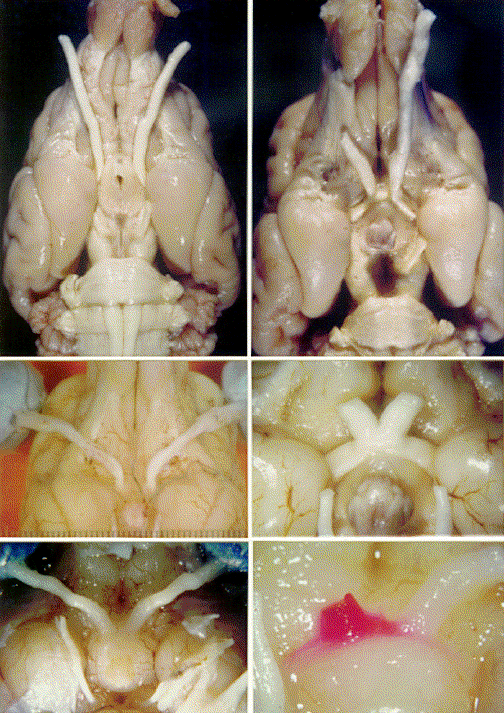
253-299 (1972). Legends Figure 1. Ventral views of Belgian sheepdog brains showing the normal
(A) and mutant achiasmatic phenotypes (case M3) (B). A. In the normal
dog, the two optic nerves meet at midline, and 80-85% of the retinal
fibers from each nerve cross midline to enter the contralateral optic
tract. B. In most mutants, the chiasm is eliminated entirely, and all
fibers extend into the ipsilateral tract. The minimum distance separated
the two retinal projections is about 5 mm. The arrowhead points to the
lamina terminalis of case M3. This structure is normally covered
ventrally by the optic chiasm. The pituitary has been removed in B,
exposing the floor of the third ventricle at the midline. Scale bar = 5
mm. Figure 2. Lamination of the LGN in normal and achiasmatic mutant
dogs. A. In this parasagittal plane, the A layer of the LGN of a normal
dog appears S-shaped, with its caudal (left) and rostral (right) parts
wrapping dorsally and ventrally, respectively. Layer A1 is separated
from layer A by a cell-sparse interlaminar zone. The magnocellular C
layer (Cm) follows the S-shape caudally and extends beneath both layers
A and A1 in the rostral, binocular portion of the nucleus. Unlike cats,
there is no interlaminar zone between layer Cm and layer A1 in the dog
(8). The parvicellular C layers (Cp) are located in the most caudal and
ventral portions of the LGN. These layers are thinner than Cm and are
separated from Cm by a narrow interlaminar zone. B. Parasagittal section
through the LGN of mutant M3. The overall shape and size of the nucleus
is similar to that of normal dogs. However, the interlaminar zone
between layers A and A1 is only present in the mid and caudal portions
of the nucleus. In the rostral LGN, layers A and A1 are tightly apposed
or fused in the mutant. Similar lamination defects may be present in the
C layers, but they are not obvious in Nissl-stained material. Cell size
does not differ from that of normal animals. C. Photomicrograph of a
coronal section through the LGN of mutant M1. Fusion of A and A1 is
evident on the medial (left) side of the nucleus. The asterisk indicates
the location of an electrolytic lesion made along penetration P6 at the
junction of A1 with Cm, and corresponds in position to the double
asterisk in Figure 3. Scale bar on each photomicrograph equals 1 mm. Figure 3. Receptive field discontinuities in the achiasmatic mutant.
These data are taken from case M1. Note the large receptive field
discontinuities (long curved arrows) along each of three penetrations
through the LGN (P6\0xD0P8). The large, nearly symmetrical shifts in
azimuth across the vertical meridian representation (VM) is larger in
the more lateral penetrations (P7 and P8). Electrolytic lesions (*) were
used to verify that the jumps occurred at the borders between layers.
Double asterisk marks a lesion along P6 that is also marked in Fig. 2C.
Penetrations P7 and P8 were so far caudal in the LGN that the electrode
initially traversed layer A1, before finally entering layer A. In
contrast, more rostral penetrations initially traversed layer A and then
passed into A1. The most lateral penetration (P5) extended through the
far peripheral representation that would represent the monocular segment
of the LGN in a normal dog. The progression of fields from high to low
elevation is normal, and is caused by the dorsoventral orientation of
geniculate layers. Small inset is a schematic plan view of the four
electrode penetrations. (HM, the approximate horizontal meridian
representation; VM, the approximate vertical meridian representation).
METHODS. We recorded from three mutant dogs ranging in age from 3
months to 1 year (M1, M2 and M3; 5\0xD022 kg). As controls, we recorded
from one behaviorally normal Belgian and three normal adult mongrels
(8-24 kg). Dogs were sedated with ketamine hydrochloride (7 mg/kg) and
xylazine (0.4 mg/kg). Anesthesia was induced with sodium pentobarbital
(15\0xD020 mg/kg). During the course of the experiments, anesthesia was
maintained by infusing sodium pentobarbital (3 mg/kg/hr). Paralytic
agents were not used. Heart rate was monitored. The eyes were protected
with contact lenses, stabilized by suturing to eye rings, and focused on
a tangent screen 57 cm from the dogs' eyes. A craniotomy was made over
the thalamus. Low-impedance tungsten microelectrodes (1 MOhm) were used
to record from visually responsive cells in the LGN. Whenever possible,
receptive fields were plotted every 100\0xD0200 um along penetrations
through the LGN, and small electrolytic lesions were made to facilitate
electrode track identification and reconstruction. At the conclusion of
the experiment, animals were given an overdose of pentobarbital
(30\0xD040 mg/kg, i.v.) and perfused transcardially with fixative. The
brain, eyes, and optic nerves were dissected free from the cranium.
Brains were sectioned at 50 um on a freezing microtome and stained with
cresyl violet. Figure 4. Simple schemes of lamination and the representation of
visual space in layers A and A1 of the carnivore LGN. The horizontal
meridian of the visual field is represented as a numbered semicircle. In
the schematics of the right LGN, the dark areas represent abnormal
representations of the ipsilateral visual field A. The pattern of
lamination, retinotopy, and visuotopy of normal dogs is comparable to
that seen in other carnivores. The area of contralateral visual space
viewed by the two eyes are aligned in precise register across layers A
and A1. B. The achiasmatic pattern in which the entire ipsilateral
hemifield (numbers on dark blocks) is represented in layer A. These
ipsilateral fields in layer A are out of register with the contralateral
fields in layer A1. C. The common pattern in the LGN of Siamese cat and
albino ferret in which lamination abnormality affects only the medial
part of layer A1. D. The pattern following prenatal or early postnatal
enucleation in carnivores (13, 19). Contralateral to the remaining eye,
layers A and A1 have formed a composite layer A/A1. Since 11 August 98
|
Neurogenetics at University of Tennessee Health Science Center
| Print Friendly | Top of Page |
Mouse Brain Library | Related Sites | Complextrait.org
